Nervesystemet er kroppens overordnede kommunikasjonsnettverk. Det samordner all aktivitet i kroppen, enten det er bevegelser, sanseopplevelser, tanker eller automatiske funksjoner som regulering av blodtrykk og tarmbevegelser. For å forstå hvordan kroppen koordinerer dette enorme spekteret av funksjoner, må vi først skille mellom de to hoveddelene av nervesystemet: det sentrale og det perifere.
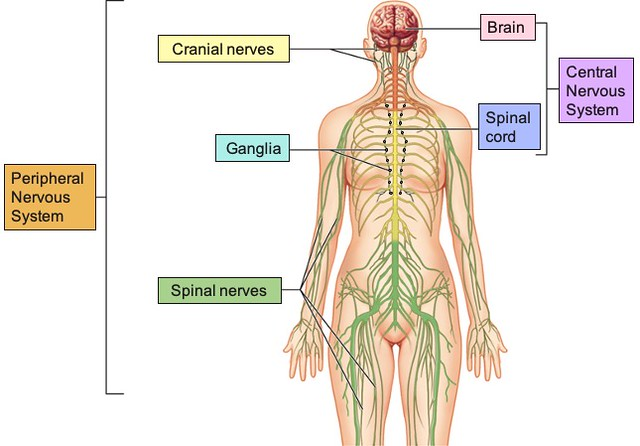
Sentralnervesystemet
Det sentrale nervesystemet (SNS) består av hjernen og ryggmargen. Hjernen fungerer som kroppens overordnede beslutningstaker og kontrollsenter, mens ryggmargen fungerer som en kommunikasjonsvei og refleksmekanisme. All bevisst og ubevisst kontroll av kroppens funksjoner har sitt utspring her.
Det perifere nervesystemet
Det perifere nervesystemet (PNS) omfatter alle nerver og nerveutløpere som ligger utenfor hjerne- og ryggmargshinnene. Det fungerer som koblingen mellom sentralnervesystemet og resten av kroppen – både huden, musklene, kjertlene og de indre organene.
Det perifere nervesystemet deles inn i to hovedkomponenter:
- Det somatiske nervesystemet – styrer viljestyrte bevegelser og formidler sanseinformasjon fra hud, ledd og muskler tilbake til sentralnervesystemet. Det er dette systemet som aktiveres når du løfter hånden for å vinke, eller kjenner at noe er kaldt.
- Det autonome nervesystemet – regulerer kroppens indre miljø og kontrollerer funksjoner som ikke er under viljens kontroll, som hjerteslag, blodtrykk, fordøyelse og pupillestørrelse. Det autonome nervesystemet deles videre inn i to komponenter med motsatt effekt:
- Det sympatiske nervesystemet, kjent som kroppens «fight or flight»-system, aktiveres i stressituasjoner. Det øker hjertefrekvensen, utvider luftveiene og mobiliserer energi slik at kroppen er klar til handling.
- Det parasympatiske nervesystemet, ofte kalt «rest and digest», dominerer i hvile og fremmer fordøyelse, energilagring og generell restitusjon.
Disse systemene jobber tett sammen for å sikre balanse i kroppens funksjoner – både i hvile og under aktivitet.
Ryggmargen og spinalnervene
Ryggmargen er en forlengelse av hjernen og utgjør den nedre delen av sentralnervesystemet. Den ligger beskyttet i ryggsøylen og strekker seg fra hjernestammen og ned til omtrent nivå L1–L2 hos voksne. Herfra fortsetter de nederste nerverøttene som en bunt av tråder, kjent som cauda equina – «hestehalen».
Oppbygning av ryggmargen
Tverrsnittet av ryggmargen viser en karakteristisk H-formet grå substans omgitt av hvit substans. Den grå substansen inneholder nervecellekropper, mens den hvite substansen består av myeliniserte aksoner som leder signaler opp og ned langs ryggmargen.
På hver side av ryggmargen går det ut et par spinalnerver – én fra høyre og én fra venstre. Hver spinalnerve består av to røtter:
- Dorsalroten (bakre rot) fører sensorisk informasjon inn til ryggmargen. Cellelegemene til disse sensoriske nevronene ligger i dorsalrotsgangliene, rett utenfor ryggmargen.
- Ventralroten (fremre rot) fører motoriske signaler fra ryggmargen og ut til kroppens muskler.
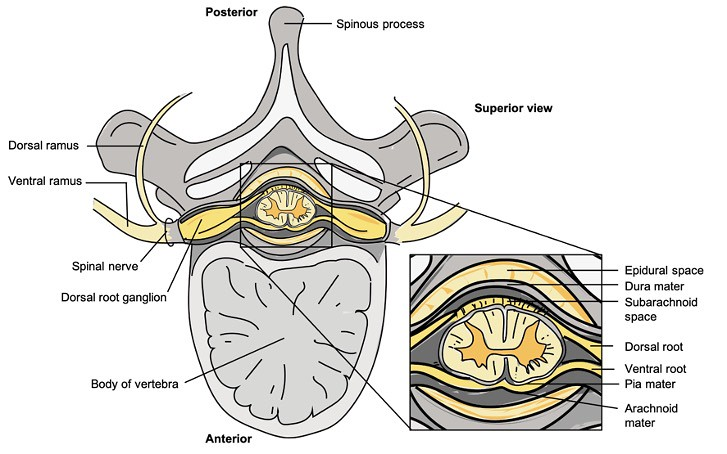
Når dorsal- og ventralroten smelter sammen, dannes en blandet spinalnerve som inneholder både sensoriske og motoriske fibre. Etter en kort fellesbane deler spinalnerven seg videre i to grener:
- Ramus dorsalis, som går til ryggens bakside og innerverer hud og dype ryggmuskler.
- Ramus ventralis, som forsyner resten av kroppen – og som danner ulike nerveplexuser, som plexus brachialis (armens nerver) og plexus lumbosacralis (bein og bekken).
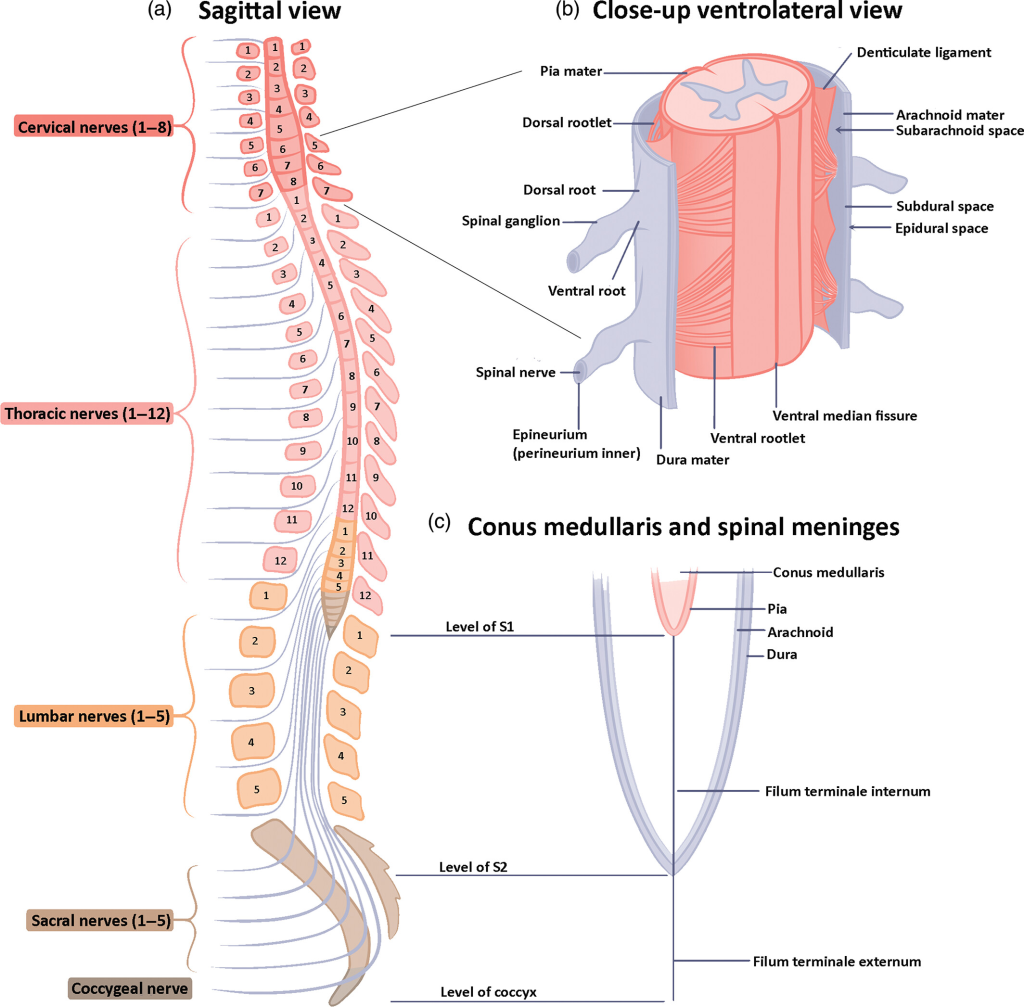
Totalt har vi 31 par spinalnerver, og disse navngis etter hvilket ryggmargssegment de springer ut fra:
- 8 cervikale (C1–C8)
- 12 torakale (T1–T12)
- 5 lumbale (L1–L5)
- 5 sakrale (S1–S5)
- 1 coccygeal
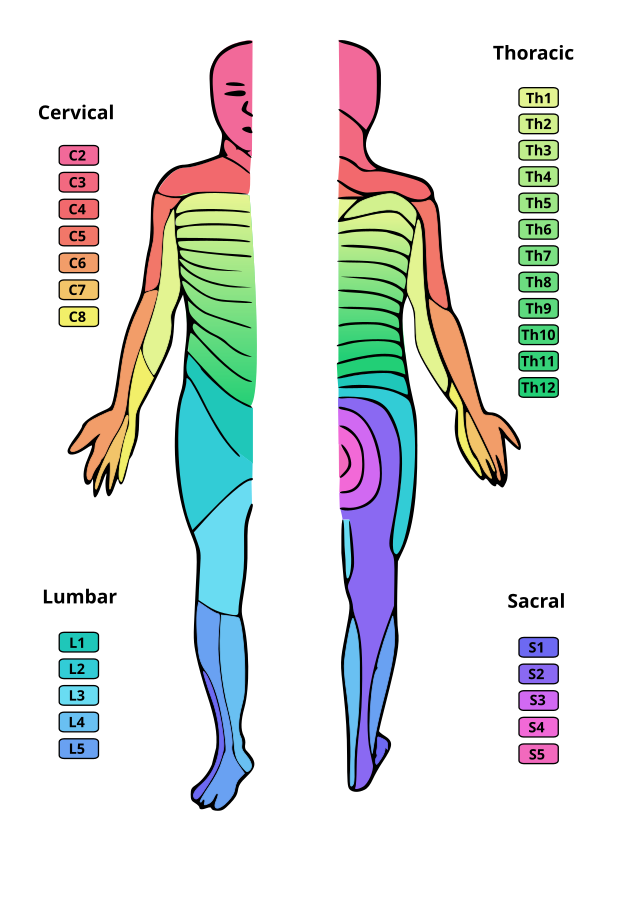
Dermatom og myotom
Hver spinalnerve dekker et spesifikt område på kroppen, både for sensorikk og motorikk:
- Et dermatom er et hudområde som innerveres av én enkelt spinalnerve. Dermatomer følger et stripemønster over kroppen og er svært viktige i diagnostikk av nerveskader. For eksempel dekker L4-dermatomet forsiden av leggen og medialsiden av foten.
- Et myotom er en gruppe muskler som innerveres av én spinalnerve. Ved å teste muskelstyrke i ulike muskelgrupper, kan man få informasjon om hvor en eventuell nerveskade sitter. For eksempel kontrollerer C5-myotomet muskler som deltoideus og biceps brachii.
Det finnes en viss grad av overlapping mellom dermatomer og myotomer, noe som gir kroppen en viss robusthet mot skader. En skade på én nerve fører derfor sjelden til fullstendig bortfall av følelse eller motorisk funksjon, men heller til delvis redusert funksjon i det aktuelle området.
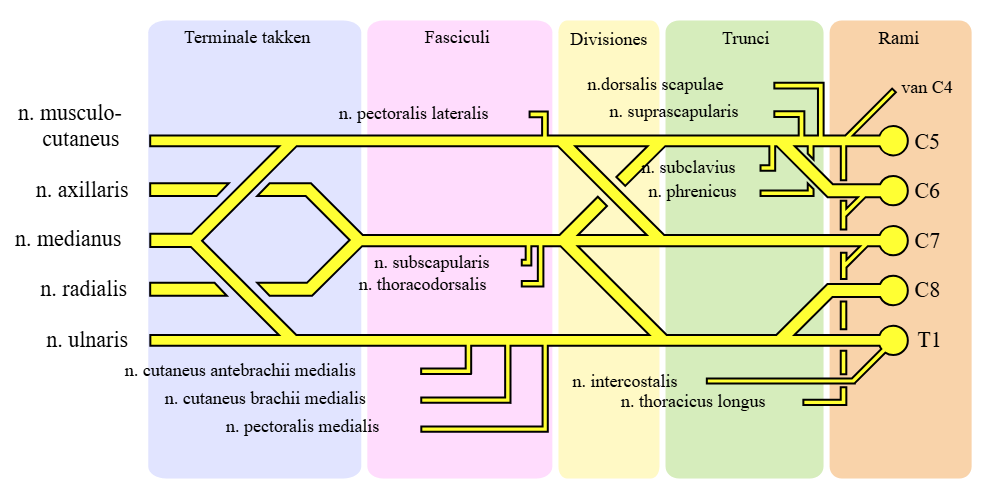
Perifere nerver og deres funksjoner
Perifere nerver er kroppens kommunikasjonskabler. De knytter sentralnervesystemet – hjernen og ryggmargen – til resten av kroppen. Disse nervene inneholder ulike typer fibre med forskjellige funksjoner, og en og samme nerve kan inneholde både motoriske, sensoriske og autonome fibre.
Hvilke signaler går i en perifer nerve?
De perifere nervene inneholder tre hovedtyper fibre:
- Motoriske fibre, som sender signaler fra ryggmargen ut til skjelettmuskulaturen, og får musklene til å trekke seg sammen.
- Sensoriske fibre, som fører informasjon fra sansereseptorer i hud, muskler og ledd inn til ryggmargen og hjernen – alt fra berøring, smerte og temperatur til propriosepsjon.
- Autonome fibre, som styrer ufrivillige funksjoner som blodstrøm, svetting og temperaturregulering i vevet.
Det betyr at én og samme nerve kan gi både bevegelse, følelse og kontrollere svette i et avgrenset område. Et godt eksempel på dette er n. medianus.
Eksempel: Nervus medianus
N. medianus er en viktig perifer nerve i armen som springer ut fra plexus brachialis. Den består av fibre fra ryggmargssegmentene C5–T1, og løper fra overarmen via underarmen og ut til hånden.
Motorisk funksjon:
Nerven styrer flere muskler i underarmen og hånden. Den gir finmotorisk kontroll til blant annet thenar-muskulaturen (tommelfingerens korte muskler), som gjør det mulig å gripe, holde og manipulere små gjenstander.
Sensorisk funksjon:
Nerven forsyner huden på den palmare (håndflate-) siden av tommel, pekefinger, langfinger og halve ringfinger. Dette er et område med høy følsomhet, og derfor svært viktig for presis berøringssans og haptisk persepsjon.
Autonom funksjon:
I tillegg til motoriske og sensoriske fibre, inneholder nerven også autonome fibre. Disse regulerer blant annet blodårenes diameter, svetteproduksjon og små muskelceller i huden (arrektorpili) i det samme området.
Klinisk betydning:
Ved skade på n. medianus, for eksempel ved karpaltunnelsyndrom, kan pasienten få både nedsatt følelse og redusert kraft i tommelens muskler. Dette skyldes at både sensoriske og motoriske fibre er påvirket. Autonom dysfunksjon kan også oppstå – som tørr hud eller temperaturforstyrrelser i hånden.
Klassifisering av perifere nervefibre
Når man studerer nervesystemet på detaljnivå, er det viktig å forstå hvordan nervefibre grupperes etter funksjon, diameter, myelinisering og ledningshastighet. Det finnes to hovedsystemer for klassifisering av perifere nervefibre: Erlanger–Gasser-systemet, som brukes for motoriske og sensoriske fibre i hud og autonome nerver, og Lloyd–Hunt-systemet, som primært brukes for sensoriske fibre fra muskel og ledd.
Erlanger–Gasser-systemet: A-, B- og C-fibre
Dette systemet benyttes hovedsakelig for:
- Somatiske motoriske fibre som styrer skjelettmuskulatur
- Sensoriske fibre fra huden (hudafferenser)
- Autonome fibre, både pre- og postganglionære
Fibre i denne inndelingen klassifiseres etter størrelse og grad av myelinisering, noe som også bestemmer hvor raskt de leder elektriske signaler.
| Fiber | Myelinisering | Diameter | Ledningshastighet | Funksjon |
|---|---|---|---|---|
| Aα | Tykkt myelinisert | 13–20 μm | 80–120 m/s | Skjelettmuskulatur (alfa-motornevroner), propriosepsjon (muskel- og senespoler) |
| Aβ | Myelinisert | 6–12 μm | 35–75 m/s | Hudreseptorer (berøring, trykk, vibrasjon) |
| Aγ | Myelinisert | 5–8 μm | 12–30 m/s | Motornevroner til muskelspoler (regulering av muskeltonus) |
| Aδ | Tynn myelinisert | 1–5 μm | 5–35 m/s | Rask, skarp smerte og temperatur |
| B | Tynn myelinisert | <3 μm | 3–15 m/s | Preganglionære autonome fibre |
| C | Umyelinisert | 0,4–1,2 μm | 0,5–2 m/s | Langsom, brennende smerte, temperatur, postganglionære autonome fibre |
A-fibrene er generelt raske, mens B-fibrene er tregere og brukes hovedsakelig i det autonome nervesystemet. C-fibrene er de tregeste og mangler myelin, men har viktige roller i både smerteformidling og autonom funksjon.
Lloyd–Hunt-systemet: Ia, Ib, II, III og IV
Dette systemet brukes utelukkende for sensoriske fibre som formidler signaler fra muskelspoler, senespoler og andre dype strukturer i muskel og ledd. Fibrene er rangert etter ledningshastighet, som henger nøye sammen med graden av myelinisering.
| Fiber | Myelinisering | Opprinnelse | Funksjon |
|---|---|---|---|
| Ia | Tykt myelinisert | Muskelspoler | Rask registrering av strekk og hastighet (strekkrefleks) |
| Ib | Tykt myelinisert | Senespoler | Måler muskelspenning – beskytter mot overbelastning |
| II | Myelinisert | Muskelspoler (statiske) | Måler lengde/stilling – propriosepsjon |
| III | Tynn myelinisert | Frie nerveender i muskel/hud | Temperatur og stikkende smerte |
| IV | Umyelinisert | Frie nerveender | Langsom, dyp smerte og kjemisk stimuli |
Ia- og Ib-fibrene er de raskeste og viktigst i reflekskontroll, spesielt i regulering av muskelspenning og koordinasjon. De langsommere III- og IV-fibrene leder informasjon om smerte og temperatur fra muskel og hud, tilsvarende Aδ- og C-fibre i Erlanger–Gasser-systemet.
Klinisk betydning
Forståelsen av disse systemene er essensiell i både klinisk nevrologi og anestesi. Ved bruk av lokalbedøvelse er det typisk at C-fibrene blokkeres først, noe som forklarer hvorfor smerteopplevelse forsvinner før berøring og motorikk. Temperaturfølelse (Aδ) forsvinner etter hvert, mens berøring og motorisk funksjon (Aβ og Aα) er mest resistente og rammes til slutt.
Tilsvarende gjelder ved kompresjon eller iskemi: de store, myeliniserte fibrene (Aα og Aβ) er mest sårbare og rammes tidlig. Dette gir kliniske symptomer som nummenhet og muskelsvakhet før smerteopplevelse svekkes. Dermed kan man ved enkel klinisk undersøkelse av sensorikk og motorikk danne seg et bilde av hvilke fibre som er påvirket – og hvor i nervesystemet problemet sitter.
Licens: CC BY SA 3.0
Hudreseptorer og sensorisk signaloverføring
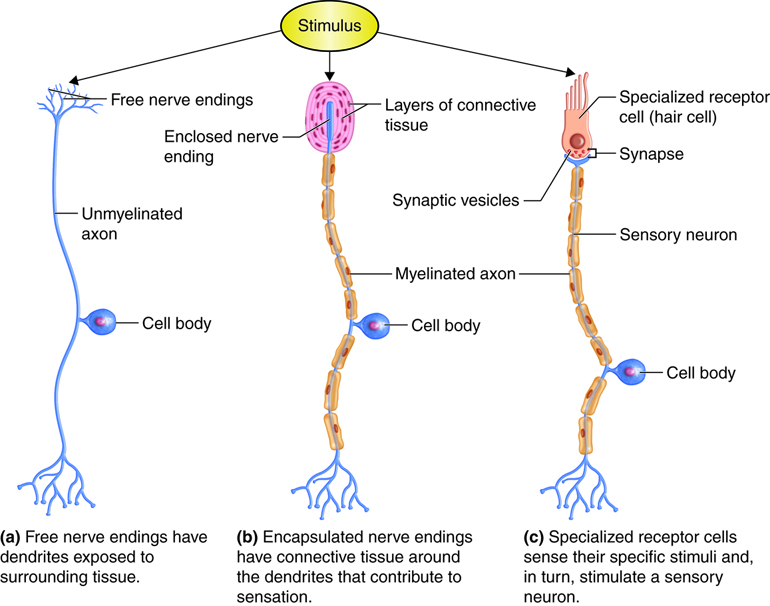
Huden er kroppens største sanseorgan. Den beskytter oss mot omgivelsene, men er også full av spesialiserte reseptorer som kontinuerlig sender informasjon til hjernen. Når vi kjenner berøring, temperatur, vibrasjon eller smerte, er det fordi reseptorer i huden aktiveres og sender elektriske signaler inn til nervesystemet. Denne signaloverføringen starter i huden, ved en reseptor.
Alt starter med en ytre påvirkning. Kanskje du berører en kald metallflate, eller noen stryker deg lett på armen. Denne påvirkningen treffer en spesialisert reseptor som reagerer på akkurat denne typen stimulus. For eksempel finnes det egne reseptorer for varme, kulde, trykk og smerte. Når reseptoren aktiveres, åpnes ionekanaler i membranen, og ioner strømmer inn eller ut av cellen. Dette skaper et elektrisk signal – et reseptorpotensial.
Hvis reseptorpotensialet er sterkt nok til å nå terskelverdien, vil det utløse et aksjonspotensial i nervefiberen som er koblet til reseptoren. Dette signalet sprer seg som en elektrisk impuls langs nerven – og det er slik nervesystemet “forstår” at noe har skjedd i huden.
Signalet følger nervefiberen innover mot ryggmargen. Der kobles det videre til neste nervecelle, og føres oppover gjennom ryggmarg og hjernestamme, til det til slutt når den somatosensoriske hjernebarken – området i hjernen som tolker følelsesinntrykk. Først da blir vi bevisst det vi har kjent.
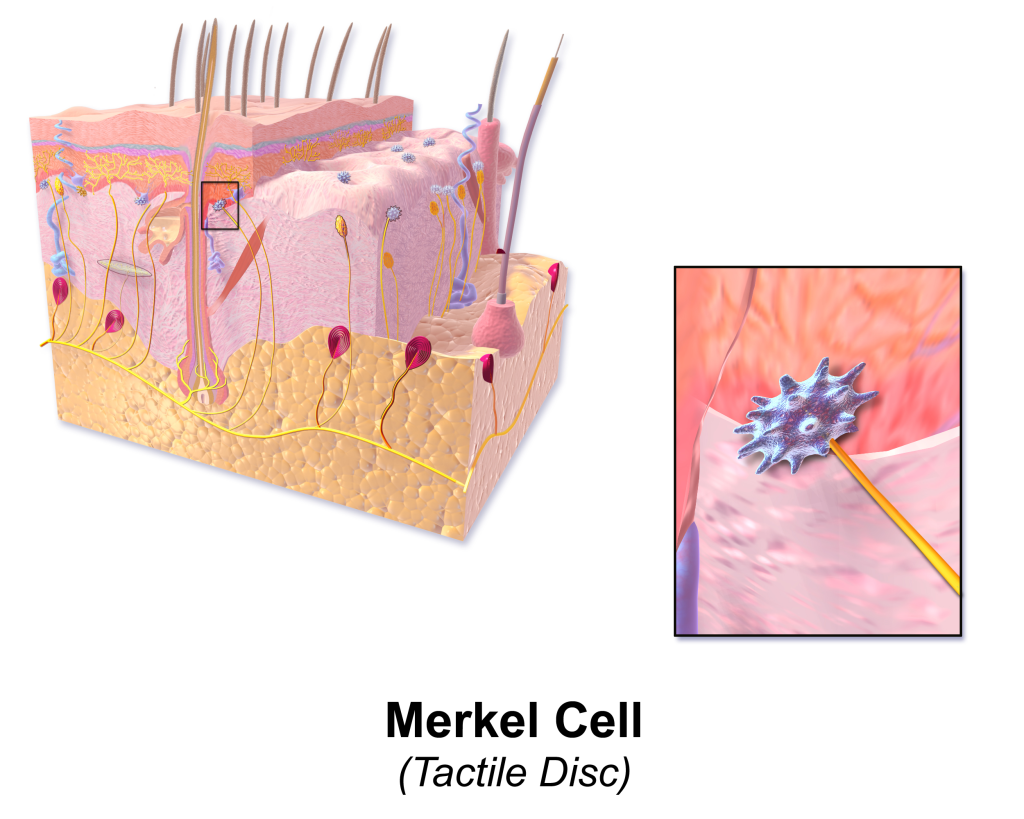
Typer kutane reseptorer
Huden inneholder flere ulike typer reseptorer, som hver er spesialisert for å fange opp én bestemt type stimuli. De fleste av disse er mekanoreseptorer – altså reseptorer som aktiveres av mekaniske endringer som trykk, vibrasjon eller strekk. Noen ligger helt overfladisk i huden og registrerer lett berøring, mens andre ligger dypere og reagerer på sterkere påvirkning.
Merkel-celler
Merkel-reseptorer ligger dypt i overhuden, tett ved basalmembranen, og finnes særlig mange steder der vi trenger høy følsomhet – som i fingertuppene. De aktiveres av lett trykk og registrerer form og tekstur på objekter vi berører. Merkel-celler er langsomt adapterende, noe som betyr at de holder seg aktive så lenge trykket vedvarer.
Meissner-legemer
Meissner-legemer er raskt adapterende reseptorer som finnes i hudens papillære lag – rett under epidermis. De er følsomme for lette berøringer og registrerer raske endringer, som når noen stryker deg på huden. De spiller en viktig rolle i finmotorikk og grep – for eksempel når du holder en blyant eller kjenner forskjellen på ulike stoffer.
Ruffini-endelser
Ruffini-reseptorer ligger dypere i huden og er spesielt følsomme for strekk og deformasjon av hud og leddkapsler. De er viktige for propriosepsjon – altså evnen til å kjenne bevegelse og posisjon i kroppen. De er langsomt adapterende og gir stabil informasjon om endringer i hudspenning.
Pacini-legemer
Pacini-legemer er de største reseptorene og ligger dypt i underhuden og i bindevev rundt ledd og sener. De aktiveres av raskt trykk og høyfrekvent vibrasjon. For eksempel er det Pacini-legemene som gjør at du kjenner mobilen vibrere, eller føler verktøyet du holder i vibrere mot huden.
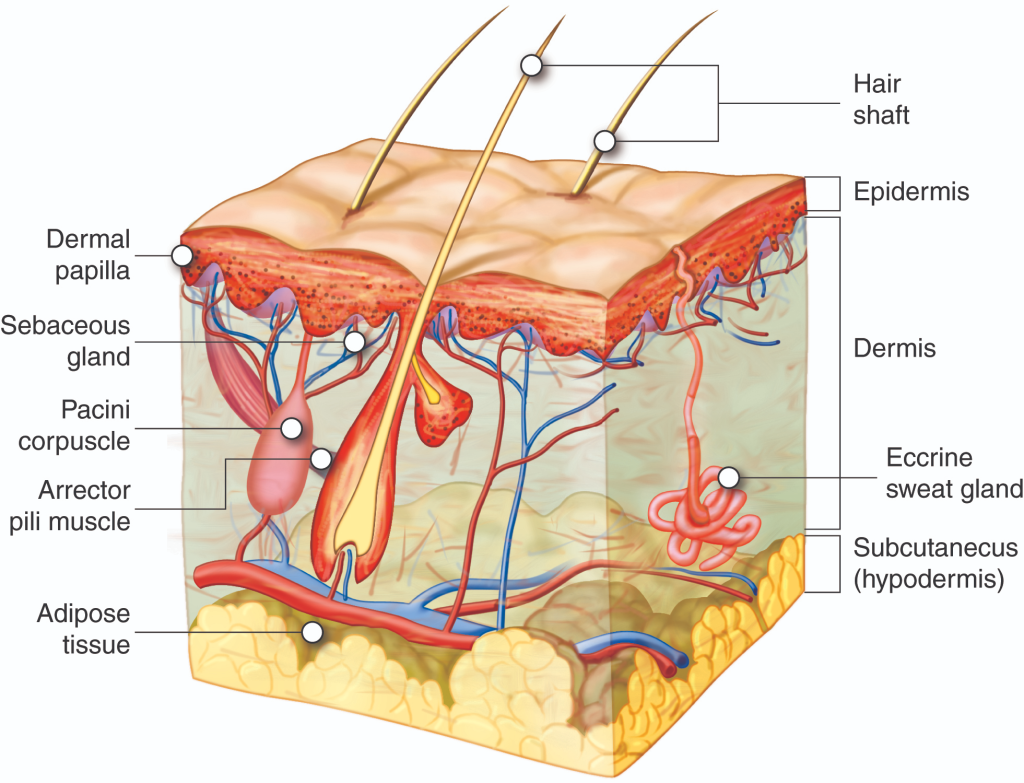
Termoreseptorer
I huden finnes også spesialiserte reseptorer for temperatur. Disse er ikke mekanoreseptorer, men ionekanaler av typen TRP (transient receptor potential). De finnes i frie nerveender og aktiveres av spesifikke temperaturintervaller.
- Kuldereseptorer (typisk Aδ-fibre) aktiveres når temperaturen faller under ca. 32 °C. De gir rask, presis informasjon.
- Varmereseptorer (typisk C-fibre) aktiveres over ca. 32 °C og sender langsommere signaler.
Mange av disse reseptorene reagerer også på kjemiske stoffer – for eksempel aktiveres kuldereseptoren TRPM8 av mentol, og varmereseptoren TRPV1 av capsaicin i chili.
Nociseptorer
Når vi snakker om smerte, er det viktig å skille mellom opplevelsen av smerte og nocisepsjon. Nocisepsjon er den fysiologiske prosessen der spesialiserte reseptorer – nociseptorer – oppdager potensielt skadelige stimuli. Disse reseptorene finnes som frie nerveender i hud, muskler og innvoller.
Det finnes flere typer nociseptorer, som reagerer på forskjellige typer trusler:
- Mekaniske: Reagerer på sterk mekanisk påvirkning, som kutt eller slag.
- Termiske: Aktiveres av skadelig varme eller kulde.
- Kjemiske: Reagerer på stoffer som frigjøres ved betennelse eller vevsskade.
Smerteformidlingen skjer via både Aδ-fibre (rask, skarp smerte) og C-fibre (langsom, brennende smerte). Kombinasjonen av disse gjør at vi først føler et stikk, og deretter en mer vedvarende smerte etter skade.
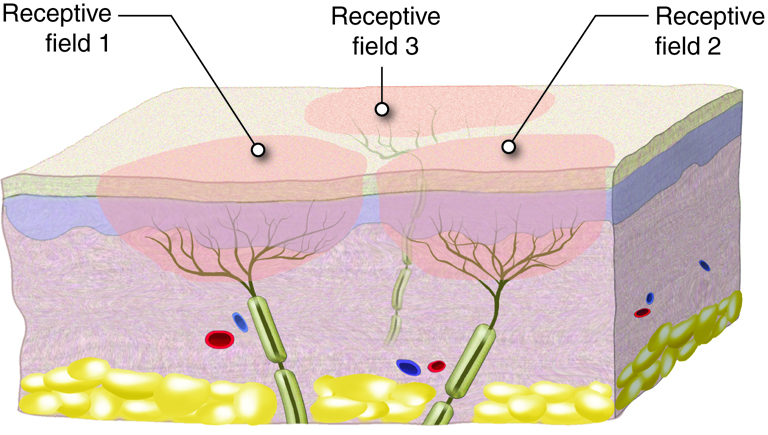
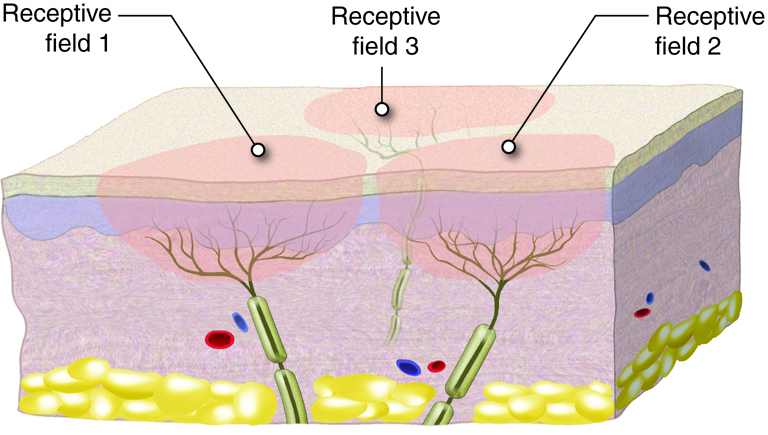{kind=link}
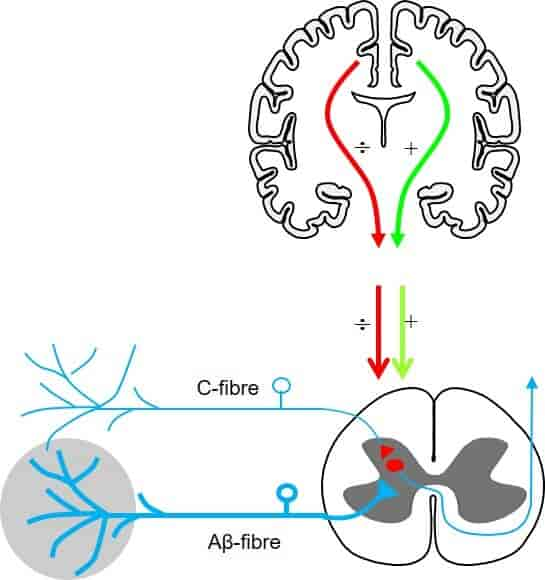
Sentral sensorisk signaloverføring – bakstrengsbanen og spinothalamiske banen
Sensorisk informasjon fra kroppen, enten det dreier seg om lett berøring, vibrasjon, smerte eller temperatur, må fraktes fra reseptorer i huden og vevet til hjernen for bevisst oppfattelse og videre bearbeiding. Denne transporten skjer gjennom to hovedsensoriske ledningsbaner i sentralnervesystemet: bakstrengsbanen (også kalt bakstreng–lemniscus-medialissystemet) og den spinothalamiske banen. Disse to systemene formidler ulike typer sensorisk informasjon og har karakteristiske anatomiske forskjeller når det gjelder både fiberbanenes plassering og hvor kryssing av midtlinjen skjer.
Bakstrengsbanen – presisjonens hovedvei
Bakstrengsbanen er ansvarlig for overføring av informasjon knyttet til berøring med høy oppløsning, vibrasjonssans og propriosepsjon (stilling- og bevegelsessans fra muskler og ledd). Denne typen sanseinformasjon er viktig for finmotorikk, gjenkjennelse av objekter ved berøring (stereognosi), og for kroppens evne til å kjenne sin egen posisjon i rommet, selv uten syn.
Signalet starter i en lavterskel mekanoreseptor, for eksempel en Merkel-celle eller en muskelspole, og ledes via en tykk myelinisert Aβ-, Ia- eller Ib-fiber til ryggmargen. Disse sensoriske nervefibrene går inn i ryggmargen via dorsalroten, men i stedet for å danne synapser der, fortsetter de direkte oppover i ryggmargens bakstrenger – fasciculus gracilis (fra ben og underekstremiteter) og fasciculus cuneatus (fra overekstremiteter og overkropp).
Første synapse skjer i nucleus gracilis og nucleus cuneatus i nedre del av medulla oblongata. Deretter krysser fibrene midtlinjen og fortsetter oppover gjennom hjernestammen som lemniscus medialis, før signalet når thalamus – hjernens sensoriske sorteringsstasjon. Her skjer en ny synapse, og tredjeordensnevronet sender signalet videre til primær somatosensorisk korteks (S1) i hjernens parietallapp, der bevisst sanseopplevelse oppstår.
Bakstrengsbanen er altså presis og rask, og viktig for sansing der nøyaktig lokalisering og detaljert informasjon er avgjørende. Den krysser midtlinjen først i hjernestammen, ikke i ryggmargen.
Den spinothalamiske banen – kroppens varslingssystem
Mens bakstrengsbanen spesialiserer seg på presis og detaljert sansing, er den spinothalamiske banen spesielt viktig for smerte, temperatur, og grovere berøring (f.eks. trykk og kløe). Denne typen informasjon er ofte knyttet til potensielle trusler og behov for rask respons, og har derfor et raskere og mer direkte opplegg.
Signalet starter her i frie nerveender – for eksempel en nociseptor eller en termoreseptor – og ledes via Aδ- eller C-fibre til ryggmargen. I motsetning til bakstrengsbanen danner disse fibrene synapse allerede i bakhornet i ryggmargen, ofte i lamina I og II (substantia gelatinosa).
Det andre ordens nevronet krysser deretter midtlinjen i ryggmargen, ofte innen ett eller to ryggmargssegmenter, og stiger deretter oppover i tractus spinothalamicus – en trakt som ligger anterolateralt i ryggmargen og hjernestammen. Når den når thalamus, skjer det en ny synapse før signalet sendes til primær sensorisk korteks (S1) for bevisst oppfattelse. Spinothalamiske fibre sender også signaler til flere andre områder i hjernen, blant annet den insulære cortex og gyrus cinguli, som er viktige for den emosjonelle og oppmerksomhetsmessige komponenten av smerte.
Et viktig poeng er at den spinothalamiske banen krysser på spinalt nivå, altså mye tidligere enn bakstrengsbanen. Det betyr at en skade på én side av ryggmargen vil gi motsidige sensoriske bortfall, men hvilke sanser som rammes, avhenger av hvilken bane som er skadet.
Klinisk betydning – en ryggmargsskades dobbeltsidige bilde
I klinikken kan man bruke kunnskapen om disse to banene til å lokalisere og tolke nevrologiske skader. Et klassisk eksempel er ved hemiseksjon av ryggmargen (Brown-Séquard-syndrom). I dette tilfellet vil man få:
- Tap av vibrasjonssans og propriosepsjon på samme side som skaden (på grunn av bakstrengsbanens ukryssede forløp i ryggmargen)
- Tap av smerte og temperatur på motsatt side (på grunn av spinothalamiske banens kryssing i ryggmargen)
Denne asymmetrien gjør det mulig å identifisere både skadeomfang og plassering ut fra en klinisk nevrologisk undersøkelse.
Motoriske signalbaner og motorisk kontroll
Motoriske signalbaner utgjør forbindelsen mellom hjernen og kroppens muskler, og muliggjør alt fra bevisste bevegelser som å løfte en kopp til automatiske justeringer av holdning og balanse. Systemet er hierarkisk og integrert, der ulike nivåer i sentralnervesystemet samarbeider for å sikre både viljestyrt kontroll og refleksbasert tilpasning.
Øvre og nedre motornevroner – en todelt kommandokjede
Motoriske baner kan forstås ut fra to hovedkomponenter: øvre motornevroner, som har cellekropp i hjernebarken eller hjernestammen, og nedre motornevroner, som har cellekropp i ryggmargen eller hjernenervekjerner og sender aksoner direkte til muskler.
Øvre motornevroner fungerer som overordnede styringsenheter som planlegger og gir ordre, mens nedre motornevroner er de utførende enhetene som aktiverer muskelfibrene. Alt somatiske muskelarbeid, enten det er å skrive, gå eller smile, er avhengig av denne todelte strukturen.
Tractus corticospinalis
Den viktigste motoriske ledningsbanen er tractus corticospinalis, også kjent som pyramidebanen. Denne starter i motorisk korteks (gyrus precentralis) og fører signaler helt ned til ryggmargens forhorn, hvor de synapser på alfa-motornevroner.
Pyramidebanen deles i to hovedgrener:
- Tractus corticospinalis lateralis: Dette er den største delen (ca. 90 %) og krysser over til motsatt side i nedre del av medulla oblongata (pyramidekrysningen). Denne delen kontrollerer de distale ekstremitetene og er avgjørende for presisjonsbevegelser, som å gripe små gjenstander eller skrive.
- Tractus corticospinalis anterior: Denne mindre delen (ca. 10 %) krysser først ved ryggmargens segmentnivå og styrer mer aksiale og proksimale muskler, som er viktige for holdning og grovere bevegelser.
Skade i tractus corticospinalis gir karakteristiske symptomer som spastisitet, hyperaktive reflekser og svakhet, spesielt i de muskler som vanligvis kontrolleres presist av hjernen.
Indirekte baner – finjustering, balanse og reflekser
I tillegg til den direkte pyramidebanen finnes det flere indirekte baner som har utgangspunkt i hjernestammen. Disse inkluderer blant annet rubrospinalis, retikulospinalis, vestibulospinalis og tectospinalis. De har sentrale roller i å regulere balanse, muskeltonus, posturale justeringer og koordinasjon av bevegelser.
Disse banene samhandler tett med cerebellum og basalgangliene, som fungerer som regulatoriske kretser. Cerebellum bidrar til koordinasjon og timing, mens basalgangliene er viktige for å starte, stoppe og velge riktige bevegelser.
Motorisk homunculus – et kart over kroppen i hjernen
I den motoriske korteks finnes det en somatotopisk organisering der ulike kroppsdeler representeres på ulike områder. Dette kalles den motoriske homunculus, et kart hvor større områder tildeles kroppsdeler med behov for presis kontroll, som fingre, lepper og ansikt.
Denne organiseringen gjør det mulig for hjernen å sende nøyaktige signaler til spesifikke muskler, og forklarer hvorfor for eksempel skader i visse deler av hjernebarken gir utfall i spesifikke kroppsdeler.
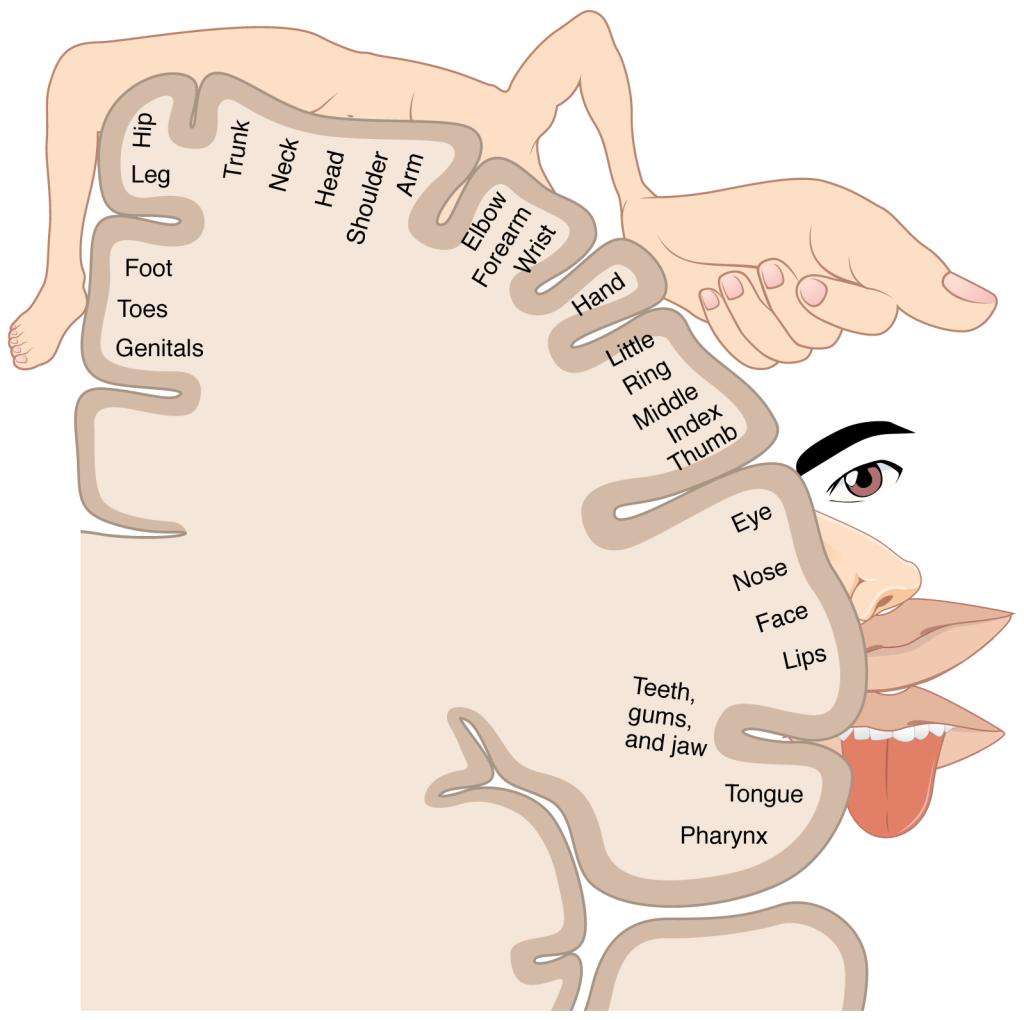
Rekruttering av motoriske enheter – justering av kraft og kontroll
Hver nedre motornevron styrer en motorisk enhet, som består av alle muskelfibrene det innerverer. For å regulere muskelkraft finnes det to hovedmekanismer:
- Frekvensmodulering – Høyere frekvens gir sterkere kraftutvikling i en enkelt enhet.
- Rekruttering – Flere motoriske enheter aktiveres etter behov. Dette skjer i henhold til «size principle», hvor små og utholdende enheter aktiveres først, mens store og kraftige enheter rekrutteres ved behov for maksimal innsats.
Denne dynamiske kontrollen gjør det mulig å tilpasse kraft og utholdenhet etter oppgaven, enten det er å holde hodet oppe i timevis eller å hoppe høyt.
Reflekser og refleksbuer
Reflekser er kroppens raskeste og mest automatiserte måter å svare på ytre stimuli. De er grunnleggende for å beskytte kroppen mot skade, opprettholde holdning og stabilitet, og regulere muskeltonus og bevegelse – helt uten at vi trenger å tenke.
Hva er en refleks?
En refleks er en automatisk og ufrivillig respons på en stimulus, formidlet via en refleksbue. Refleksen skjer uten at signalet nødvendigvis må opp til bevisst bearbeiding i hjernen først. I stedet bearbeides det i ryggmargen eller hjernestammen, noe som gir raskere respons.
Den klassiske refleksbuen – kroppens snarvei
En refleksbue består av fem grunnleggende komponenter:
- Reseptor – Registrerer endringer i omgivelsene, som strekk i en muskel eller smerte fra en nål.
- Afferent ledd (sensorisk fiber) – Leder informasjon inn til ryggmargen.
- Reflekssenter – I ryggmargen bearbeides signalet og det bestemmes om en respons skal utløses.
- Efferent ledd (motorisk fiber) – Leder signalet ut til muskel eller kjertel.
- Effektor – Utfører responsen, for eksempel en muskelsammentrekning.
Strekkerefleksen – kroppens balansevakt
Den myotatiske refleksen, også kjent som strekkerefleksen, er kroppens viktigste verktøy for å opprettholde muskeltonus og leddstabilitet. Den er monosynaptisk – det vil si at signalet fra sansecellen går rett til motornevronet uten mellomstasjon.
Hvordan fungerer det?
Når en muskel plutselig strekkes, som ved slag på patellasenen (knehammerrefleks), registrerer muskelspolen denne raske strekkingen. Muskelspolen sender signaler via Ia-afferente fibre til ryggmargen. Der danner de en direkte synapse med alfa-motornevronet, som aktiverer samme muskel og får den til å trekke seg sammen.
Samtidig skjer det også en resiprok inhibisjon via internevroner, som hemmer antagonisten – altså muskelen som jobber mot bevegelsen – slik at refleksen blir mer effektiv og målrettet.
Kliniske eksempler på strekkerefleks:
- Bicepsrefleksen (C5–C6)
- Tricepsrefleksen (C6–C7)
- Patellarrefleksen (L2–L4)
- Akillessenerefleksen (S1–S2)
Fravær eller overaktivitet i disse refleksene kan tyde på skade i ryggmargen eller perifere nerver.
Fleksjonsrefleksen – kroppens forsvar
Den polysynaptiske fleksjonsrefleksen utløses typisk av smerte, og beskytter kroppen ved å trekke bort den utsatte kroppsdelen. Den er et klassisk eksempel på en refleks som går via flere interneuroner.
Eksempel:
Du tråkker på en spiss gjenstand – nociseptorer i fotsålen aktiveres og sender signal via Aδ-fibre til ryggmargen. Der aktiveres flere internevroner som stimulerer fleksormuskulatur i benet og samtidig hemmer ekstensorene. Resultatet er at du trekker benet unna.
Samtidig skjer det ofte en krysset ekstensjonsrefleks, der den motsatte fotens strekkemuskulatur aktiveres for å opprettholde balanse. Dette viser at refleksene våre er del av større, koordinert motorikk – ikke bare enkle reaksjoner.
Ia-inhibitoriske internevroner og resiprok inhibisjon
Når en muskel aktiveres, er det avgjørende at dens antagonist ikke jobber imot. Dette reguleres av Ia-inhibitoriske internevroner. De får signal fra muskelspolens Ia-fibre og hemmer alfa-motornevronet til antagonisten. Dette skjer automatisk ved bevegelser og er en forutsetning for jevn, kontrollert motorikk.
Renshawceller – intern brems på motoraktivitet
Renshawceller er en spesiell type inhibitoriske internevroner som aktiveres av kollateraler fra et alfa-motornevron og gir negativ tilbakekobling. Det betyr at når motornevronet fyrer, sendes samtidig et signal til en Renshawcelle som demper aktiviteten til det samme nevronet og eventuelt nærliggende motornevroner.
Dette hindrer overaktivering og bidrar til stabilisering av motoriske signaler, spesielt under repetitive bevegelser som gange. Skade på Renshawceller eller deres signalvei kan føre til spastisitet og ukoordinerte bevegelser.
📚 Anki-kort
Obs, tomt! Kommer etterhvert <3
📝 Eksamensoppgaver
Obs, tomt! Kommer etterhvert <3
👨⚕️ Klinisk case
Obs, tomt! Kommer etterhvert <3
❓ Test deg selv
Obs, tomt! Kommer etterhvert <3